@ilya-antipin Да, почитал енто дело. Получается, канадцы в этом исследовании сначала «забраковали» предыдущую модель строения матричной оболочки ВИЧ Альфадли и Барклиса, которые в 2007 и в 2009 годах в условиях in vitro при помощи кристаллографического анализа показали, что матричные белки ВИЧ с хвостами остатков миристиновой кислоты на плоской фосфолипидной мембране в растворе, во-первых, собираются в тримеры, а во-вторых, из этих тримеров образуют правильные шестиугольники (все стороны равны и углы по 120 градусов).
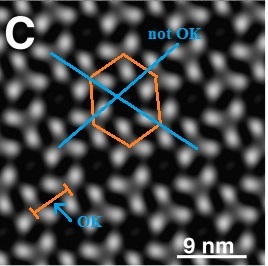
Точнее, про тримеры канадцы оставили, а вот то, что сфера матрикса может быть образована правильными шестиугольниками забраковали. Забраковали на основе формулы Эйлера и здравого смысла, который подсказывает, что любая вершина многогранника, образованная какими-либо плоскими фигурами, на плоскости должна иметь сумму углов в данной вершине меньше 360 градусов (в нашем случае 3 правильных шестиугольника на плоскости в точке их соединения образуют 3 угла по 120 градусов, что равняется 360 градусам и, следовательно, это невозможно).
На картинках ниже это видно, что при придании сферической формы тримеры в составе шестиугольников при следовании от экватора к полюсам (направление «сжатия» сферы показано стрелками) наезжают друг на друга и, тем самым, теряется правильная форма шестиугольников ближе к полюсам, вплоть до сильного перекрытия тримерами друг друга.

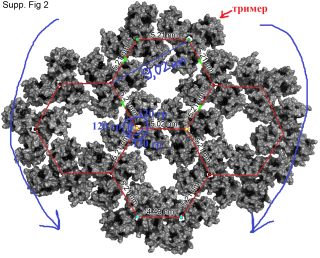
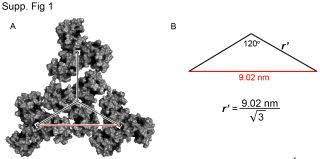
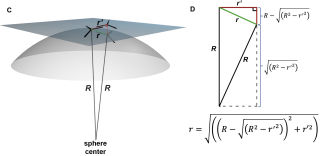
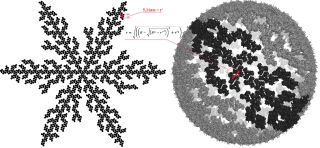
Также канадцы использовали полученное Альфадли значение 9,02 нм, которое равняется расстоянию на плоскости между центрами масс определенных тримеров в шестиугольнике и использовали это расстояние для расчета расстояния между соседними тримерами и получили 5,21нм, затем вывели формулу для корректировки этого расстояния при переносе на сферическую поверхность в зависимости от радиуса данной сферы матричной оболочки ВИЧ (колеблется где-то от 62,5 до 72,5 нм). Затем с использованием математической модели (использование матрицы вращения и компьютерных программ) вывели определенную решетку/структуру расположения тримеров в образованной ими же сфере. Она получилась в виде 6 долек/лепестков с центральными прямыми хребтами в каждой и на северном полюсе, например, плотная, а на южном – неплотная. А вдоль самих лепестков и между лепестками из тримеров расстояния между тримерами по сфере очень различаются, образуя в том числе большие пустоты.
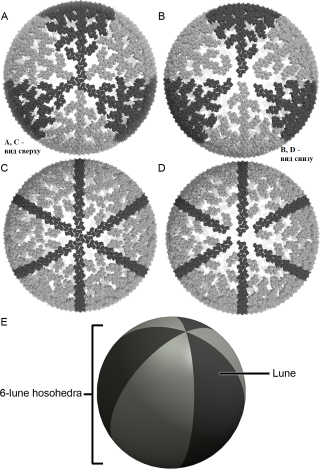
И далее авторы на основе этой модели предположили механизм проникновения капсида и слияния матриксной оболочки с клеткой человека как раз южным полюсом с большой пустотой за счет размыкания лепестков (словно медуза).
Некоторые мысли:
- Получается, из плоскостной структуры Альфадли из правильных шестиугольников саму идею шестиугольников канадцы посчитали некорректной (вполне обоснованно), а вот расстояния между образующими эти шестиугольники тримерами (5,21 нм, полученное на основании расстояния 9,02 нм) они посчитали корректным и на основании этого расстояния на плоскости выдвинули свою теоретическую гипотезу строения матриксной сферы в виде лепестков. Немного странно?
- Н у и интуитивно, как-то не верится в такую структуру матричной сферы вируса с такими неравномерно распределенными пустотами между лепестками. А гликопротеины gp120 и gp41 тоже получается неравномерно распределены по шарику вируса? А слияние с лимфоцитами происходит только, если вирион прибьется/развернется/прокатится строго южным полюсом к лимфоциту? Как-то слишком умно для туповатого ВИЧ.