Биологи впервые отследили процесс проникновения ВИЧ в клетки человека
-
Участник @gremlin написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
а) обмена между разновирионными геномными РНК и разновирионными ревертазами (в том числе при рекомбинациях) во время обратных транскрипций;
Ретровирь псевдодиплоидный – два полных генома в РНК несет, но эти два полных генома транслируются в одну провирусную ДНК, и вот тут и происходит рекомбинация (два сценария, см. PMID: 30307534, и с гомозиготами просто тут результат не отследить никак. Интересно, считал ли кто сколько тех самых гомозигот… к чему я. А так ли важно, если у тебя две РНК, меняться промеж вирусов? Разнообразие и в двух соседних РНК будет, если будет, то же (не принципиально иным), на первый взгляд, как и с РНК из другого вириона. Нет? Не уверен тут.
-
Участник @gremlin написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
через поры ядра
Коразтыкенет. Не через поры. В том и фишка этого ядерного эндоцитоза, поры по боку.
It was found that after entering the cell, the viral particles moved along the microtubules and selectively gathered at the microtubule organization center (MTOC), leading the nearby nuclear envelope (NE) to undergo deformation, invagination and restoration to form a nuclear vesicle in which the viral particles were wrapped; then, the inner membrane of the nuclear vesicle ruptured to release HIV-1 into the nucleus. This phenomenon is similar to cell endocytosis and therefore called the “nuclear endocytosis-like pathway”. Factors involving in the process were preliminarily investigated. This discovery expands our understanding of the complexity of HIV-1 nuclear entry, which may provide new insights to HIV-1 virology. The molecular details and viral biology of the mechanism need further elucidation.
-
Участник @ilya-antipin написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
А так ли важно, если у тебя две РНК, меняться промеж вирусов? Разнообразие и в двух соседних РНК будет, если будет, то же (не принципиально иным), на первый взгляд, как и с РНК из другого вириона. Нет? Не уверен тут.
Однозначно, считаю, что разнообразие было бы намного выше при существовании такого «разновирионнорекомбинантного» механизма. И намного опасней с точки зрения возникновения лекарственно-устойчивых штаммов.
Гетерозиготы ВИЧ могут появляться только при условии наличия сразу не менее 2-х разных провирусных ДНК в геноме инфицированной клетки. Причем выход гетерозигот при, например, 2-х провирусных ДНК в среднем будет около 50% в сравнении с гомозиготами.
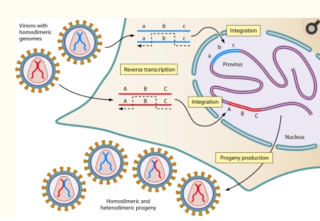
А вот таких клеток (с 2-мя и более провирусами в геноме) в организме, что недавно инфицированных, что хронически инфицированных пациентов не более 10, максимум 15% [1], [2], [3].
Предположим, у нас есть 10 клеток с разными провирусными ДНК в составе. Пусть согласно исследованиям 9 клеток несут один провирус, а 1 клетка несет 2 провируса в геноме. Пусть все эти 11 провирусных ДНК имеют каждая по одной какой-либо разной мутации устойчивости, в том числе к разным классам препаратов.
Тогда при их реактивации в более-менее один временной период эта армада, состоящая из приблизительно 19/20 части гомозиготных и 1/20 части (50% от 1/10) гетерозиготных вирионов рассеется по организму и клеткам. И уже на этой стадии, т.к. разные вирионы, по-любому, будут залетать в одни и те же клетки, в случае возможности «межвирионной» рекомбинации в цитоплазме начнут со страшной силой смешиваться и образовывать всевозможные мутанты (ревертаза прыгает с шаблона на шаблон РНК от 5 до 10 раз), в том числе мультирезистентные. И тем более, вичевые ревертазы не будут ограничиваться скачками только «туда-обратно» меж 2-х РНК, а будут скакать, как угорелые, между множеством шаблонов вирусных РНК «туда-сюда-туды-сюды-обратно», типа того. Т.е., практически, каждая вновь инфицированная клетка будет обладать би-, либо полимутантными провирусными ДНК.
А вот, если ревертазе ВИЧ можно скакать только внутри капсида, то потенциальный (но еще не готовый) рекомбинант сможет образоваться только из 10% вновь инфицированных клеток, где будет «впаяно» по 2 и более провируса на клеточную ДНК. И то рекомбинант от этих клеток будет образовываться где-то в 50% дочерней популяции и к тому же будет содержать одновременно лишь не более 2-х мутаций устойчивости (т.к. РНК смешиваться в один вирион будут только по 2-е, хоть из 2-х, хоть из 5-и одновременно «впаяных» в геном клетки разных провирусов), максимум 3-х мутаций устойчивости (в случае, если один из провирусов в геноме клетки происходит от атаки 1/20 гетерозиготной части вирионов).
И так далее по циклу преимущество в диверсификации «межвирионнорекомбинантной» популяции по сравнению с «внутрикапсиднорекомбинантной» популяцией будет только наращиваться.Оно и понятно, если бы такая «межвирионная» рекомбинация была возможна, то мы бы имели на сегодня кучу полирезистентных штаммов ВИЧ у каждого пациента.
Но конечно, технически, наверняка, было бы сложно ревертазам в цитоплазме на расстояниях скакать с РНК на РНК, нежели компактно внутри капсида. Но это уже другой вопрос
-
Участник @ilya-antipin написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
Коразтыкенет. Не через поры. В том и фишка этого ядерного эндоцитоза, поры по боку.
Да, конечно, там же все черным по белому было написано.
“nuclear endocytosis-like pathway”
Бес дернул меня наускоре накинуть про импорт в ядро.))
А по поводу экспорта из ядра, как думаете?
У меня такая версия: я так, подозреваю, что на гекса(пента)мере капсида p24 где-то есть аминокислотная метка (ядерный экспортный сигнал NES) для экспортина и Ran-GTP. Подозреваю, что эта «метка» замаскирована, когда капсид в сборе, а вот когда разбирается, то она оголяется. Вот только до каких структурных элементов разбирается капсид? До мономеров, до гексамеров, или до конгломератов гексамеров? И, следовательно, при разборке капсида до какого структурного элемента, метка (NES) оголяется и становится видимой для экспортин- Ran-GTP комплекса?
Типа того:
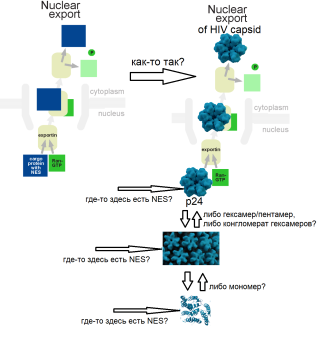
??? -
Участник @gremlin написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
про импорт в ядро.))
А по поводу экспорта из ядра, как думаете?Может я ошибаюсь, но нужны секторальные санкции на импорт. И чтобы запрещенка не проникла даже через Беларусь. А если проникнет, то давить ее бульдозером .
-
Участник @gremlin написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
Гетерозиготы ВИЧ могут появляться только при условии наличия сразу не менее 2-х разных провирусных ДНК в геноме инфицированной клетки.
А точность обратной транскриптазы мы здесь игнорируем? Сколько там… 5-10 на 9000 помню, 1/1700 замен/добавлений/делеций помню цифру… Ну, или тут почитать можно.
-
@васьвась а лучше давить тракторами не запрещенку, а тех, кто ее ел.
-
Участник @gremlin написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
А по поводу экспорта из ядра, как думаете?
Стараюсь не думать об этом )) Я в такого рода вопросах все же склонен ждать, когда большие пацаны определятся, все разжуют для моего уровня, и вот тогда кушать уже и других кормить.
-
Участник @gremlin написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
Однозначно, считаю, что разнообразие было бы намного выше при существовании такого «разновирионнорекомбинантного» механизма.
Интуитивно – да. Но я погонял эту мысль так и сяк, и что-то стал сомневаться. Пока не готов обсуждать, думать нужно, рисовать и прикидывать.
-
Участник @ilya-antipin написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
А точность обратной транскриптазы мы здесь игнорируем?
Ни в коем случае. Без ошибок обратной транскриптазы рекомбинация была бы почти бессмысленной с точки зрения образования различных мутантов (в том числе мультирезистентных) и гетерогенности популяции ВИЧ. Почти, потому что и сама рекомбинация в виде прыжков обратной транскриптазы с шаблона РНК на шаблон может служить причиной делеций и вставок. Но какое отношение точность обратной транскриптазы имеет к самому непосредственному образованию гетерозиготных вирионов, кроме того, что она основная изначальная причина существования разных геномов ВИЧ- материалов для гетерозигот?
Но я, наверное, уловил Вашу мысль. Вы, вероятно, имели в виду, что невысокая точность обратной транскриптазы ВИЧ может нивелировать значение рекомбинации в развитии генетического разнообразия ВИЧ и, как следствие, в возникновении полирезистентных штаммов ВИЧ.
Сколько там… 5-10 на 9000 помню, 1/1700 замен/добавлений/делеций помню цифру…
Это, по-моему, старенькие данные.
Если даже взять Вашу же свежую ссылку (интересная кстати статья о влиянии на конечную оценку точности ревертазы ВИЧ точности транскрипции шаблона РНК для дальнейшего его использования под обратную транскрипцию ревертазой ВИЧ).Ну, или тут почитать можно.
то там получается «differences in fidelity between the studied RTs with error rates ranging within 2.5 × 10−5 and 3.5 × 10−5», т.е. от 1 ошибки на 40000 нуклеотидов до 1 ошибки на 28571 нуклеотидов.
Правда, это в условиях in vitro, но в in vivo вот ребята в 2015г. тоже проводили расчет и у них получилось «This reveals an extremely high mutation rate of (4.1 ± 1.7) × 10−3 per base per cell.»
Но, затем выясняется, что «We show that the HIV-1 reverse transcriptase contributes only 2% of mutations, whereas 98% result from editing by host cytidine deaminases of the A3 family.» Т.е. те же порядки получаются, что и в исследовании по Вашей ссылке in vitro, а именно = (8,2 ± 3,4)×10-5 на нуклеотид, т.е. где-то в среднем 1 ошибка ревертазы на 12000 нуклеотидов.Но, в принципе, эта точность обратной транскриптазы не так уж сильно и важна для нашей дискуссии.
Даже если мы возьмем 1 – 2, или чуть больше ошибок на геном (геном ВИЧ около 10000 нуклеотидов), то все равно вероятность того, что ревертаза совершит 2 ошибки на геном в нужном нам месте нужным образом (с заменой исходной дикой аминокислоты на нужную нам) чрезвычайно низка. А вот при рекомбинации вероятность совместить эти 2 мутации (пусть из-за предварительных ошибок ОТ) в одном будущем провирусе гораздо выше, с учетом того, что ревертаза прыгает с шаблона на шаблон РНК по 5 – 10 раз за обратную транскрипцию.
Например, возьмем 2 мутации устойчивости к разным классам препаратов M184V в гене обратной транскриптазы и R263K в гене интегразы. Между этими позициями в геноме 1915 нуклеотидов. И представить себе, что ОТ ошибется именно в 2-х этих позициях нужным образом довольно сложно (пусть и последовательным наложением мутаций), а вот представить, что при вышеуказанной частоте прыжков с одного мутантного шаблона на другой мутантный шаблон при рекомбинации ей перескочить на интервале в 2000 (если точнее 1915) нуклеотидов из 10000 вполне вероятно (можно даже скакануть на этом интервале не один раз, главное нечетное число). И тут бы мы сходу получили полирезистентный штамм, а если бы существовала межвирионная рекомбинация, то достаточно быстро бы получили такой результат.
А вот при внутрикапсидной рекомбинации нам бы пришлось долго ждать пока 2 наших мономутанта, вышедшие из разных клеток, войдут в одну и ту же клетку и именно оба успешно встроят свои ДНК в один-единственный геном этой клетки (таких клеток, напомню, около 10%). И потом именно гетерозиготные отпрыски из этой клетки выживут, успешно рекомбинируют нужным образом и успешно встроят свой рекомбинантный геном в клетку-мишень. -
Тут осенило, Илья Игоревич. Возможно, в своём предыдущем сообщении я не совсем верно интерпретировал Ваш вопрос.))
@Ilya-Antipin:А точность обратной транскриптазы мы здесь игнорируем?
Вы, возможно, имели в виду ровно вот это в моем тезисе:
@Gremlin:Гетерозиготы ВИЧ могут появляться ТОЛЬКО при условии наличия сразу не менее 2-х разных провирусных ДНК в геноме инфицированной клетки.
Тогда мой ответ был бы значительно короче)):
@Gremlin:Гетерозиготы ВИЧ могут появляться только при условии наличия сразу не менее 2-х РАЗНЫХ провирусных ДНК в геноме инфицированной клетки.
Т.е. ни в коем случае не игнорировал ошибки ОТ при обр. тр-ции.
-
@gremlin проблема еще и в том, что основная масса исследований у нас основана на не очень точных данных по оценки интеграций. QVOA тут не валиден (дорого, нужно реактивировать все провирусы)… далее считали копии, но не убеждались в их целостности – тоже плохая дата. Мы к 2020 толком не знаем большую дату по числу провирусов, способных реактивировать и выделять новые вирусы, не говоря о том, как это соотносится с множественными интеграциями, а к ним еще вопросы есть и не только по целостности, но и по параметрам реактивации для разных сайтов интеграции. Мы знаем, что сайты не случайны и не равны. А как? Не очень. 80% интеграций происходит в 40% генома, но интроны и экзоны представлены вполне себе в сайтах тех. При этом оценки 13 и 16 годов – 98% дефектных провирусов. Вообще непонятно, что там с математикой общей тогда. Роль персистирующих клонов и роль провируса в управлении клональной экспансии, размеры клонов в миллионы клеток, при очень низких параметрах активированных… это вжжж неспроста. И явно что-то сильно отличается в начале и финале заболевания, а как? А в латентноф фазе опять же. А АРВТ… дохрена вопросов, и ответы на часть из них тольо множат вопросы. Но это я в сторону отошел уже.
Это, по-моему, старенькие данные.
Очень. Я знаю. Потому с лениниским прищуром к ним. И вообще все эти новые данные сильно меняют картину мира, и я бы предпочел подождать 2-3 года, прежде чем пытаться начать дальше смотреть этот сериал.
Вот простой вопрос: если мутация устойчивости находится в дефектном провирусе, но дефект не накрывает большой фрагмент, ген пусть целиком, условно, Nef там какой дефектный, но ген с той самой мутацией или мутациями другой, и где-то тут есть и целостная интеграция, то что дальше? Потерян ли полностью доступ к мутациям? Вся магия сплайсинга где, когда, из откуда…
Меня уже давно подмывает написать текст с заголовком «Наташа, проснись, мы все переписали»… но как-то пока не уложилось на уровне синопсиа даже.
-
@ilya-antipin в один день вышли сразу два интересных исследования. Первое - определенные комбинации из 8-10 белков позволяют надежно идентифицировать клетки резервуара, способные вызвать активную инфекцию.
https://www.amfar.org/HIV-Reservoir-Cells/
Мне кажется это серьезный шаг вперёд. Ранее не было известных маркеров, позволяющих отделить инфицированные клетки от здоровых.
Второе - для активизации вируса необходима комбинация из нескольких LRA. Одни эффективны для активизации резервуаров вируса в крови, вторые - в тканях, третьи - в лимфатических узлах.
https://www.amfar.org/Blood-and-Gut-Respond-Differently-to-Virus-Activators/
-
Участник @авб написал в Биологи впервые отследили процесс проникновения ВИЧ в клетки человека:
Мне кажется это серьезный шаг вперёд.
Очень непонятно как это знание применить. Устроить аутоиммунную атаку на них? Ну, такое… Сделать вектор, который будет делать действие по наличию этих белков – не умеем пока и мутно тут все. Хорошее знание, но, боюсь, что пока бесполезное, тем более, что речь о кучке фенотипических кластеров, т.е. ничего универсального тут не сварить по определению. Для исследований эти сортировки занятная штука, там применимость может дать что-то косвенно.
Второе – вообще не могу оценить значимость, тканевые различия и так были понятны, ну, вот так оно.
-
Молекулярное моделирование показало, как ВИЧ уклоняется от иммунной системы
Ученые смогли смоделировать процесс заражения ВИЧ. Эксперты надеются, что открытие поможет в разработке новых методов лечения. Работа опубликована в Proceedings of the National Academy of Sciences.Вирусы ВИЧ-1 — это эволюционный результат редкой успешной передачи вируса иммунодефицита обезьян (ВИО) человеку. Поскольку люди не восприимчивы к ВИО, ученые задались вопросом, как пандемическая группа ВИЧ способна уклоняться от иммунной системы человека. Существуют также другие, непандемические штаммы (группы ВИЧ-N, ВИЧ-O и ВИЧ-P), они очень редки и были обнаружены лишь у нескольких человек в Западной Африке.
Особое внимание эксперты уделяли TRIM5α — белку, который содержится в клетках большинства приматов и эффективен против различных инфекций и ретровирусов.
TRIM5α связывается с капсидом — оболочкой, в которой заключен вирусный геном. Активность этого белка частично зависит от циклофилина А (CYPA), другого белка, обладающего каталитическими свойствами, то есть способного ускорять определенные реакции. Ученые решили разобраться, как действует этот механизм.
Чтобы запустить моделирование, для начала эксперты создали структурную модель капсида ВИЧ-1 с помощью белка циклофилина А, которая затем использовалась для создания других комплексов ВИЧ-N, ВИЧ-O и ВИЧ-P. Для сравнения исследователи также моделировали конверсию в капсиде без циклофилина А.
Всего эксперты создали 16 различных систем. С их помощью они смогли детально рассмотреть, какие пути проходит вирус, прежде чем проникнуть в клетку. Эксперимент показал, что скорость реакции, катализируемой белком циклофилином А, может играть решающую роль в защите ВИЧ-1.
То есть белок циклофилин А слегка изменяет капсид — оболочку, в которой заключен вирусный геном. Эти преобразования, как считают ученые, оказывают серьезное влияние на вирусную инфекцию.
Именно связывание циклофилина А обеспечивает защиту человеческого вируса ВИЧ-1 от антивирусного белка TRIM5α.
Таким образом, исследование выявило новое слабое место ВИЧ-1. Теперь эксперты будут работать над тем, чтобы создать лекарства, подавляющие связь циклофилина А с вирусом в клетке.